
Acoustic Physiology in Mosquitoes
- 1Graduate School of Science, Nagoya University, Nagoya 464-8602, Japan
- 2Institute for Advanced Research, Nagoya University, Nagoya 464-8602, Japan
- 3Ear Institute, University College London, London WC1X 8EE, United Kingdom
- 4The Francis Crick Institute, London NW1 1AT, United Kingdom
- ↵5Correspondence: joerg.albert{at}ucl.ac.uk
Abstract
The acoustic physiology of mosquitoes is perhaps the most complex within the entire insect class. Past research has uncovered several of its—sometimes stunningly unconventional—principles, but many mysteries remain. Their solution necessitates a concerted transdisciplinary effort to successfully link the neuroanatomical and biophysical properties of mosquito flagellar ears to the behavioral ecology of entire mosquito populations. Neuroanatomically, mosquito ears can rival those of humans in both complexity and sheer size. The approximately 16,000 auditory hair cells within the human organ of Corti, for example, are matched by the approximately 16,000 auditory neurons in the Johnston's organ of a male Anopheles mosquito. Both human and mosquito ears receive very extensive efferent innervation, which modulates their function in ways that are as yet poorly understood. Different populations of neuronal and nonneuronal cell types divide the labor of the mosquito ear amongst themselves. Yet, what exactly this labor is, and how it is achieved, is at best vaguely known. For the majority of mosquitoes, biologically relevant sounds are inextricably linked to their flight tones. Either these flight tones are (directly) the sounds of interest or they contribute (indirectly) to the production of audible sound through a process called nonlinear distortion. Finally, male ears can generate tones themselves: The generation of an internal “phantom copy” of a female flight tone (or self-sustained oscillation) is believed to aid the male hearing process. Here, we introduce protocols that target the mosquitoes’ auditory neuroanatomy, electrophysiology, and behavior to help shed light on some of these issues.
HEARING IS A KEY SENSE FOR MOSQUITOES
The sense of hearing is key to mosquito survival; without it, the mid-air copulation between male and female could not take place (Fig. 1A). Males identify—and locate—a female mating partner by listening out for her faint flight tones (Fig. 1B). Within the (acoustically and spatially) crowded environment of mosquito mating swarms (Downes 1969), where hundreds or thousands of males often compete for a few dozen females, this is a challenging sensory task. For a mosquito male, reproductive fitness might thus be approximated as acoustic fitness; the (positive) selection for genes that improve hearing, as well as the (negative) selection against genes that cause deafness, can be expected to have shaped the evolution of male mosquitoes.
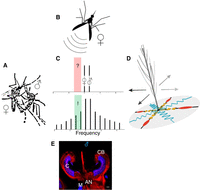
The acoustic ecology of mosquitoes. (A) Mosquitoes copulate mid-flight, following a precopulatory pursuit (often taking place in large mating swarms) during which the male chases the female. (B) Males use the females’ flight tones to identify females within the swarm and to track them during the phonotactic chase. (C) (Top) The mosquitoes’ flight tones (bars) are nearly inaudible to both sexes, as they fall outside the mosquitoes’ hearing range (red highlighted area); (bottom) when male and female flight tones mix nonlinearly within the mosquitoes’ ears, however, they produce audible distortion tones (additional bars), which fall partly within the mosquito hearing range (green highlighted area). (D) Audibility thus arises from an interaction between (male and female) flight tones and the mosquito flagellar ear (see schematic drawing). Understanding the biophysical operation of the mosquito flagellar ear is key to understanding mosquito hearing and auditory behavior. (E) Within the Johnston's organ (JO), different populations of ciliated neurons (neurons red, cilia blue), and nonneuronal cell types, coalesce to mediate, and modulate, the process of mosquito hearing. The antennal nerve carries both afferent and efferent nerve fibers, which transmit, or modulate, sensory signals, respectively. (CB) Cell body, (C) cilia, (AN) antennal nerve, (M) muscle.
Despite their crucial importance (without their sense of hearing, males could not execute the phonotactic chase of a female mating partner) and seeming simplicity (the only relevant sounds are the pure tone-like “flight tones”), the mechanisms of mosquito hearing have retained many of their secrets to this day. The assumption of simplicity has proven deceptive.
A BRIEF BREAKDOWN OF MOSQUITO HEARING
On the receiver side, the mosquitoes’ flagellar ears convert airborne vibrations (such as those associated with a flight tone) into neuro-electrical signals, which are then propagated to the mosquito brain. The flagellar shaft is the actual sound receiver: it acts like an inverted pendulum that is set into motion by friction with the surrounding air (Albert and Kozlov 2016). The greater the velocity of the air particles, the larger the flagellar oscillation. The flagellum's plane of oscillation depends on the direction of the velocity vector, and thus the location of the sound source. To boost sound-induced motion, the flagellar receivers of male mosquitoes possess arrays of fibrillar extensions (or “fibrillae”), which increase the flagellum's effective acoustic surface. In Anopheline mosquitoes these fibrillae canalso be collapsed. Mosquito ears have an exquisite sensitivity: In males, angular displacements of <1 mdeg (corresponding to flagellar tip displacements of <20 nm) (Su et al. 2018) are enough to excite the auditory neurons of the Johnston's organ (JO), the mosquitoes’ “inner ear.”
Mosquitoes, especially males, should thus be well-adapted to detect the faint sounds produced by the wingbeats of a nearby flying mate. However, testing the responses of their ears to naturally occurring flight tone frequencies tells a different story. Mosquito flight tones change with ambient temperatures (Villarreal et al. 2017). Under laboratory conditions (20°C–22°C), females emit flight tones of ∼400 Hz and males emit flight tones of ∼600 Hz. In both sexes, however, these tones fail to elicit responses in the auditory neurons of JO. Instead, the male JO shows a response plateau centered on frequencies of ∼200 Hz (ranging from ∼150 Hz to ∼250 Hz; see Somers 2022 and highlighted areas in Fig. 1C). No mosquito sound emissions in this frequency range have been reported. The solution to this puzzling mismatch is provided by an essential property of mosquito (and other) ears. As a direct consequence of their mode of operation, sound receivers display characteristic nonlinearities (e.g., gating compliances). The receivers’ nonlinearities, in turn, introduce distortions into their response to external sound. Even when stimulated with two pure tones only (such as the mosquitoes’ flight tones; see Fig. 1C, top), the receiver produces mathematically predictable distortion products (DPs) (Julicher et al. 2001), some of which will fall into the sensitivity range of the mosquitoes’ auditory nerve (Fig. 1C, bottom). The nonlinear mixing of two—in themselves inaudible—flight tones thus can create an audible distortion tone within the mosquito ear (Gibson et al. 2010; Somers et al. 2022). DPs are, however, at least two orders of magnitude smaller than the original flight tones, and thus the exquisite displacement sensitivity of the flagellar ear is crucial to their detection.
AUDIBILITY IS A COMPLEX, MULTIVARIATE PROBLEM IN MOSQUITOES
Two likewise immediate, and far-reaching, conclusions can be drawn from the relations sketched out above: (i) The audibility of females is a key prerequisite for the males’ precopulatory chase and thus for reproduction in mosquitoes in general; and (ii) audibility between mosquitoes is a nontrivial, interrelational problem, as it does not simply arise from a match between an emitted signal (e.g., a female flight tone) and a receiver's properties (e.g., a male ear). Uniquely among all other reported hearing systems, audibility in mosquitoes also depends on sounds generated (and possibly controlled) by the receiver (Fig. 1D): For a male to be able to hear a given female, he must himself fly with the correct wingbeat frequency (and thus the correct flight tone), which generates audible distortions when mixing nonlinearly with the female's flight tone in his own ear. Studies have started to shed light on these issues, exploring the mutual relation, and possible optimization, of male and female flight tones within the context of the mating swarm. However, many known (and most certainly many unknown) unknowns remain. The biggest mystery here is arguably the role of the self-sustained oscillations (SSOs) specific to male ears. SSOs represent self-generated flagellar oscillations, which closely mimic those induced by female flight tones. It has been speculated that SSOs act as internal amplifiers of female flight tones, but their specific contributions and ecological relevance—and cellular origin (Fig. 1E)—are still unclear.
FROM BIOPHYSICS TO VECTOR CONTROL
From what we do know, direct lines can be drawn from the in-depth study of mosquito auditory biology to vector control efforts. First, hearing is a key sensory modality that guides male reproductive behavior within the mating swarm. The mating swarm has been identified as a prime target for future vector control efforts (Sawadogo et al. 2017), and current programs aiming at the release of genetically modified mosquitoes have almost exclusively concentrated on males (as the nonbiting sex). A better quantitative understanding of mosquito hearing—and audibility—thus carries the dual promise of (i) providing novel molecular targets and mechanistic routes for mosquito population control efforts and—perhaps even more importantly—(ii) supporting ongoing release programs through improved predictive models of expected reproductive interactions (and compatibility) between populations of genetically modified and native mosquitoes (Su et al. 2020). One simple way that this could be achieved is to “acoustically match” mutant and wild-type mosquitoes in order to optimize their respective mating interactions. This discussion and the three associated protocols have been written in an effort to promote auditory research in mosquitoes on the cellular (see Protocol: Immunohistochemical Staining of the Mosquito Ear [Andrés and Albert 2022]), biophysical (see Protocol: Electrophysiological Measurements of Compound Action Potential Responses from the Antennal Nerve in Response to Stimulation [Su and Albert 2022], and behavioral (see Protocol: Recording and Extraction of Mosquito Flight Tones [Georgiades and Albert 2022a], and Protocol: Mosquito Phonotaxis Assay [Georgiades and Albert 2022b]) levels.
ACKNOWLEDGMENTS
Our work benefited from funding through the European Research Council (ERC) under the Horizon research and innovation programme (grant agreement no. 648709, to J.T.A.), a pump-priming award from the BBSRC Vector Borne Disease (VBD) Network ANTI-VeC (AV/PP/0028/1, to J.T.A., Sarah Moore, and M.P.S.), a BBSRC project grant (BB/V007866/1 to J.T.A.), and a UCL Global Challenges Research Fund (GCRF) small grant (to J.T.A.).
Footnotes
-
From the Mosquitoes collection, edited by Laura B. Duvall and Benjamin J. Matthews.








